Régulation de la transcription par l’ARN polymérase II au niveau de la phase d’élongation
La transcription des gènes par l’ARN polymérase II peut être divisée en plusieurs phases, notamment la préinitiation, l’initiation ou amorçage, l’élongation et la terminaison. Si les deux premières phases sont évidemment très importantes pour le contrôle de l’expression de beaucoup de gènes, ce n’est que récemment que l’on s’est rendu compte que la transition entre initiation et élongation est importante pour la modulation de l’expression de près de 30% des gènes transcrits par l’ARN polymerase II. En effet, dans beaucoup de cas cette polymérase ralenti et s’arrête même complètement (on parle de pause) après avoir synthétisé les premiers nucléotides d’un ARN messager. Le passage à une enzyme plus efficace pour poursuivre l’élongation de l’ARN nécessite des modifications post-traductionnelles de la plus grande sous-unité de l’ARN polymérase II. Cette sous-unité contient dans sa partie C-terminale de nombreuses copies d’un hexapeptide qui peut être phosphorylé au niveau de sérines en position 2 et 5 de ces répétitions. La phosphorylation des sérines 2 est particulièrement importante pour rendre la polymérase plus efficace. Cette phosphorylation est effectuée par un facteur appelé p-TEFb qui est constitué d’une kinase, Cdk9, et d’un co-facteur appelé cycline qui peut être soit la cycline T soit la cycline K.
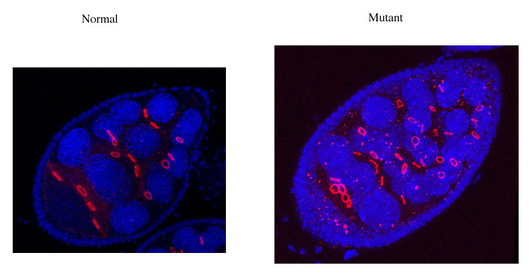
Mon laboratoire combine des approches génétiques et moléculaires chez Drosophila melanogaster pour analyser in vivo la fonction des complexes p-TEFb. Nous avons généré des transgènes qui permettent une expression contrôlée d’une kinase Cdk9 inactive qui entre en compétition avec la kinase normale. Cette approche de mutation dominante négative nous a permis de décrire les effets d’une réduction de l’activité p-TEFb dans différents tissus de la drosophile. Cela permet de décrire l’importance de p-TEFb lors de la prolifération et de la différenciation cellulaire. Ce travail est une étape dans l’identification de gènes qui sont particulièrement fortement contrôlés au niveau de la transition entre l’initiation et l’élongation (relachement de polymerase en pause). Nous avons aussi créé des mutations et des différents transgènes pour les cyclines. Nous avons pû montrer que la cycline T et la cycline K sont toutes deux indispensables et que les deux types de complexes p-TEFb ont des activités différentes.